{Readers:
Larry Gall, Peabody Museum, Yale University, New Haven, CT 06511.
E. Richard Hoebeke, Dept. Entomology, Cornell U., Ithaca, NY 14853.
Phil Kingsley, Otis Methods Development Center, USDA-APHIS, Otis, MA 02542.
Ellen Prager, Division of Biochemistry and Molecular Biology,
U. California, Berkeley, CA 94720.
Corresponding Address: Joseph G. Kunkel
Biology Department
University of Massachusetts
Amherst, MA 01003
Telephone #: (413) 335-6017
E-Mail: joe@bio.umass.edu
Differences in both yolk proteins and locations of eleven forewing venation core landmarks in Catocala (Lepidoptera: Noctuidae) behave neutrally during evolution. Yolk proteins are to be digested by embryos; composition counts, not amino acid sequence. While shape of wing surface is clearly important for flight, positions of vein branch points are less important. Venation can be substantially variant within species without compromising flight. The non-Darwinian immunological and morphological clocks we have described have similarly even tempos. Random walks of multidimensional traits have led to statistical irreversibility, a confirmation of Dollo's Law. Neutral traits can evolve in any direction; progress and retreat have little meaning in such situations. The similar distances computed between several pairs of Catocala using either anti-yolk sera or core wing landmarks suggest that either approach is valid in studying the evolution of this group. While the immunological and planar geometric approach agree with the classical alpha-taxonomic approach of comparing genitalia, a more interesting result is that they are promising as metrics for ordering higher taxonomic relationships. Evolutionary progress can be measured upon a backdrop of these clock-like traits but such progress is more likely to concern unpredictable introduction of novel uses of proteins and wings rather than metronomic neutral changes in structural details of the clocks.
KEYWORDS: immunological cross-reaction, planar geometry, Dollo's Law, non-Darwinian evolution, tree-building.
Using numerical methods to evaluate morphological and molecular differences between living species to establish phylogenetic relationships is a well-established practice (Sokal, 1961; Sokal and Sneath, 1963). However, that has not eliminated heated debate over regularity of molecular and morphological clocks and the appropriateness of using particular traits or metrics (Wilson et al., 1977, 1987; Radinsky, 1978; Carlson et al., 1978; Wood, 1983; Benjamin et al., 1984; Beverley and Wilson, 1984, 1985; Lewin, 1988; Felsenstein, 1991).
The concept of neutral morphological traits is relatively novel since most distinct morphology is thought to be functional unless it is in the process of disappearing due to disuse. Even in the case of atrophy due to disuse the trait is not thought to be neutral because benefit is derived from saving on an investment. This same apparent enigma was encountered in neutral protein and nucleic acid evolution but has been thoroughly rationalized: In nucleic acids, third base changes are often neutral when they do not change the amino acid encoded. These and amino acid changes in proteins which do not affect the activity or function of an enzyme are also candidates for neutral mutations, so-called non-Darwinian evolution (King and Jukes, 1969). The fibrinopeptide A and B regions of the fibrinogen polypeptide accept mutations at close to neutral frequency and as a result exhibit one of the most rapid protein evolutionary rates recorded (Dickerson, 1972). In other proteins the extent of residues which are neutral to change may be more restricted, with histone H2A being at the other extreme of accepting next to no neutral mutations. The extent of neutral mutations determines the overall rate of a protein's evolutionary rate of sequence change.
By analogy then, a morphological mutation which changes a structure but does not adversely affect the function of that structure could be categorized as a neutral morphological mutation and would be expected to change at some regular rate, in a random walk manner, as long as its degree of change does not threaten the function of some surrounding functional structure. In the absence of reliable time scale information, evolutionary biologists have attempted to use internal controls and comparisons to validate a particular trait's usefulness as a clock. We attempt this approach for wing morphology evolution in the underwing moths.
The evolution of vitellogenin, Vg, the major yolk protein precursor in insects, has been described as largely neutral and rapid relative to other serum proteins (Hagedorn & Kunkel, 1969; Kunkel & Nordin, 1985) based on immunological cross-reactions in cockroaches and underwing moths (Johnson, 1974; Kunkel et al., 1976). In another insect group, the family Saturniidae, broader cross-reactivity across generic boundaries is exhibited by Vg (Telfer & Williams, 1953). It is clear, however, that Vg is more rapidly evolving than other serum proteins such as hexamerins, which cross-react throughout orders of insects (Kunkel & Nordin , 1985; Telfer & Kunkel, 1991). We accept these data at face value, suggesting that they speak to the relative ages of the taxa involved. The present paper in particular attempts to validate neutral wing venation changes by demonstrating a parallelism in divergent evolution of morphological (wing vein landmarks) and biochemical (yolk protein) evolution. While wing structure might be considered a highly selected trait (Tilyard, 1919; Wootton, 1990, 1992; Johnson and Walter, 1978), we present evidence that a central core of wing venation landmarks behaves neutrally within and between species. We internally compare wing venation differences and yolk protein cross-reactivity with parallel but incomplete distance matrices (using only two reference species) and finally use a metric on wing venation differences to compute a complete morphological distance matrix with which we build an informative phenetic tree.
The practical significance of validating a neutral morphological trait is that it provides a simple method in which every taxon can be used as a reference such that a complete reciprocal distance matrix can be easily computed. In contrast, to obtain a complete distance matrix using an immunological test of difference is a daunting task, requiring an antibody to each taxon considered. The heuristic value of our example may lead to a new approach to searching for other neutral morphological traits.
Materials and Methods
Animals. -- Specimens for protein analysis were collected by M. E. Johnson, C. Kellogg and T. D. Sargent in the Franklin and Hampshire Counties of central Massachusetts. Specimens for wing venation analysis were obtained from the research collection at the University of Massachusetts curated by T. D. Sargent. Origins of specimens involved in wing venation analysis are given in Table I. The taxonomic names of species are given according to Hodges and coworkers (1983).
Protein extractions.-- Eggs were obtained from gravid females by dissecting oviducts plus ovaries from abdomens and extracting the eggs with Buffer A (0.2 N Na Cl, 0.01 N Na2 HPO4, pH 7.5). The homogenate was centrifuged (10,000 rpm, IEC B-20, 15 min 4°C) and the supernatant saved and dialysed twice against Buffer A. The yolk extract was stored frozen at -20°C.
Antiserum production.-- Antisera against the yolk proteins of two species within Section A of the lepidopteran genus Catocala (C. retecta and C. habilis) were raised in New Zealand white rabbits (Kunkel, 1987). The antisera were made female-specific by adsorbing with extracts of whole conspecific adult male moths in Buffer A. The specificity of the antisera to yolk extracts and lack of reactivity to male extracts was checked with the Oudin test. No discernible reactions of male extracts were detectable at the lowest dilutions of the adsorbed antisera.
Immunological procedures.-- The adsorbed female specific antisera were used for quantitative comparative serology in a double-diffusion-in-agar approach initiated by Butler and Leone (1968). The reactivity of each antiserum was measured toward its homologous antigen as a reference using a full precipitin titration curve technique (Boyden, 1964) specially adapted to the agar gel medium. Ouchterlony double diffusion was performed on slides with wells cut with 14-gauge cannulation needles. Seven wells were cut in 1" x 3" glass slides coated with 2 ml of 1% agarose in Buffer A using a template with center-to-center well placement of 6 mm in a linear array on the central axis of the slide. These wells each contained 3.2 µl of volume. Wells 2, 4 and 6 were filled with Ab and wells 1, 3, 5 ,7 were filled with Ag. Diffusion was allowed to occur for 24 hours at 4°C, after which the wells were flushed with Buffer A and the slides washed for 24 hours in two changes of buffer A to remove unreacted protein. The slides were then dried down by capillarity of overlying layers of filter paper and toweling for 2 hours and rinsed in distilled water to remove residual salts prior to drying. The air dried slides were stained with Thiazine Red R in 50:50 methanol, 7% acetic acid and rinsed twice in 7% acetic acid for 10 min each.
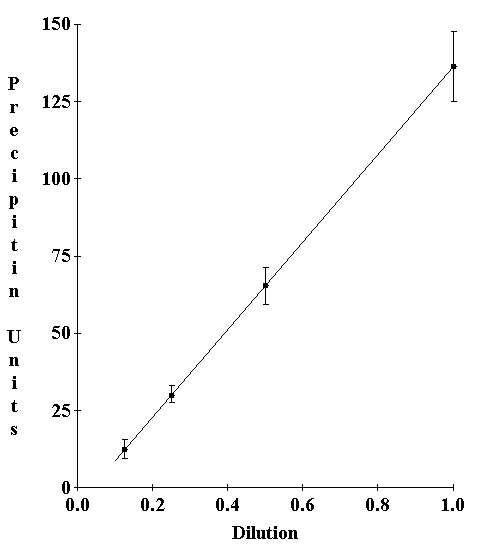
Full titration curves in liquid use a fixed antiserum concentration and measure the amount of precipitin formed as antigen is titrated from antigen excess through equivalence to antibody excess (Kunkel, 1987). This full titration curve approach requires impossibly high antigen titers to achieve the antigen excess portion of the curve in agar. To estimate this part of the curve a titration of the antibody to the highest antigen concentration was carried out. The precipitin reactant from this region of the titration curve was multiplied by the dilution factor to obtain the equivalent precipitate at full antiserum strength. Test titrations on model-antigen antibody reactions confirmed that serial dilution of both the antigen and antiserum resulted in a corresponding serial diminution of the resultant precipitin (Fig. 1).
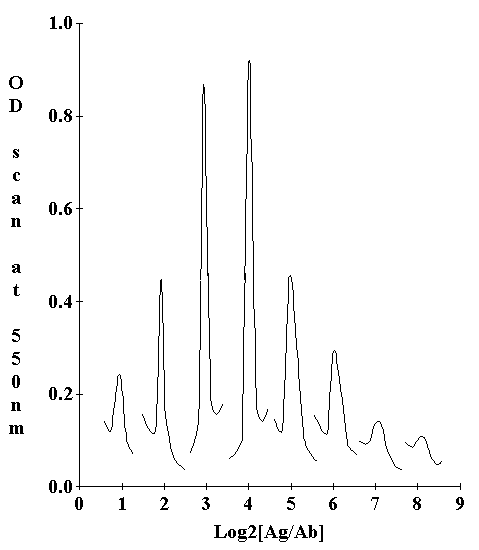
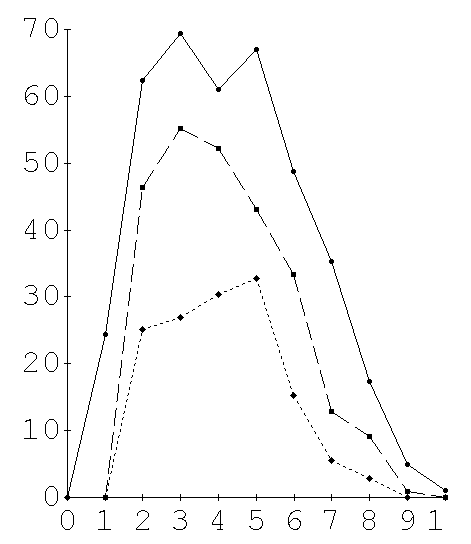
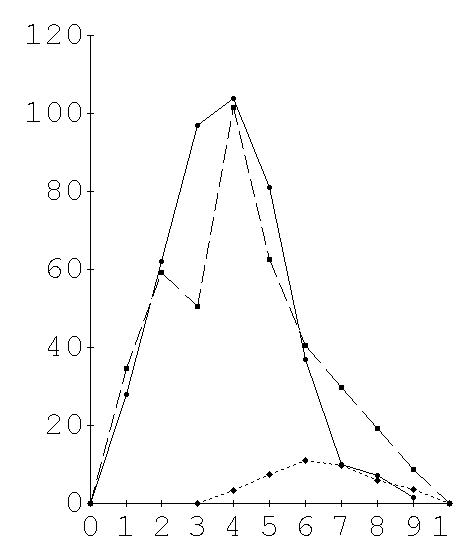
The precipitin amounts were estimated by scanning the stained slides in a Gilford spectrophotometer with a scanning attachment (Fig. 2). Six individual double diffusion tests were scanned for each Ag:Ab dilution combination. Between 6 and 10 dilution combinations were needed to achieve a full titration series depending on the antigenic divergence of the test species (Fig. 3A, 3B). The planimetric area under each curve was used as an estimate of the precipitate at that Ag:Ab dilution. A standard curve in which both Ag and Ab are serially diluted at their equivalence ratio demonstrated that the technique is quantitative and linear (Fig. 1). The observed reactivity (O) of a given species yolk extract to an antiserum was taken as the sum of all the average integrated areas for each dilution of Ag; a sum of the variances at the individual dilutions was used to estimate the variance of the full titration curve. The observed reactivity, O, for each non-homologous antigen was measured as a proportion of the expected maximum reactivity (H) exhibited by the homologous Ag. A dissimilarity coefficient, D, was computed,
Thus antigenic identity would score 0 in dissimilarity while antigenic divergence from identity would score some fraction of 1 up to 1 for a non-reacting antigen.
Wing preparation for morphological study.-- Wings were optically cleared and prepared for photography in a series of steps: (1) wet briefly in alcohol, (2) decolorize in 50-100% household bleach, (3) rinse in distilled water, (4) soak in distilled water 10 minutes, (5) dehydrate in absolute alcohol (ethyl or isopropyl), (6) degas under vacuum from a rotary vane pump, (7) arrange on acetate film on a glass plate and sandwich with alcohol and a second acetate film, expressing any bubbles and crushing the basal wing structures to create a flat sandwich, (8) apply a second glass plate and clamp the glasses together, (9) store a video image, and (10) dismount and store applied to one acetate sheet.
Wing photography.-- The wings sandwiched between glass were placed on a light box on a photographic stand and a video camera (Dage CCD-72A) fit with a 50mm macro lens was used to image the wing on a RGB monitor. A millimeter rule was imaged at the same time to provide calibration. Images were grabbed by a Matrox PIP-640 (square pixel) video digitizer board controlled by Microsoft-C software written by JGK taking advantage of the PIP-EZ library of functions for the Matrox board. Software was written which allowed image contrast to be maximized using the background, gain and contrast controls of the Dage camera controller prior to grabbing the image. The objective of this phase was to obtain the best image for all of the relevant landmarks. Images were saved onto a high-capacity disc drive over an Ethernet connection. After landmark data were extracted, the images were compressed using the Unix utility, compress, and archived to 3/4" tape at 1600 bpi.
Data extraction and storage.-- Software was written which allowed X,Y coordinates of a wing image to be stored and retrieved onto the wing image involved for verification. Landmark X and Y coordinates comprised the primary data. Outline type data, enhancements of Siegel (1981), were comprised of a series of sequential X,Y coordinates defining a continuous outline of some specimen structure. These enhancements could be specified to help orient an investigator when the landmark type data were viewed without an accompanying image. Outline data are not included in the statistical comparison of structures. Calibration data consisted of coordinates in the X-dimension of up to 10 adjacent marks of a millimeter rule. A log of the wing and specimen identification and the time its data were extracted was kept. Calibration, outline and landmark type data were stored in a common binary database file where type, X and Y coordinates and time of extraction comprise a record. The data records for any time period could be recalled onto the appropriate wing image.
Planar geometry.-- The basic algorithm used for analysis was that of Siegel (Siegel, 1981, 1982; Siegel & Benson, 1982), which was translated from FORTRAN code into Turbo Pascal by JGK. This method uses a rotation, translation and isometric scaling of one set of landmarks onto a second homologous set chosen as a model. Both the least-square and robust regression methods of Siegel (1982) were tested for utility in different phases of the analysis. In general the simple least-square approach was found most useful in the present context of differences within and between closely related species. The basic pairwise fitting of Siegel was enhanced with a within-population averaging routine which computes an average of n sets of homologous landmarks to form an average model. It accomplishes this by choosing one specimen to be a provisional model and uses the least-square approach to fit all other members of the population to that provisional model. Then the average location of a landmark with a confidence ellipse about it was recomputed by refitting all the specimens to the averaged model using the least-square regression fit; the size of the average model was scaled to equal the average size of the population since the original fit scales all data to the provisional model.
Residual differences between the p average landmarks of two populations, A and B, were combined into a single metric on the difference between A and B in two ways, an average (avg) residual distance and a root-mean-square (rms) residual distance:
and
where Ai-Bi represents the geometric distance between the ith X,Y coordinates of A and B.
Pairwise distances, avg or rms, between individual population average models were calculated using the least-square or robust regression approach (Siegel, 1982) creating a distance matrix to which a Fitch-Margoliash (F-M) tree building algorithm was applied (Fitch and Margoliash, 1967). The rms residual distance matrix was found overall to perform better in tree building than the avg residual distance matrix.
Multivariate Analysis of Dispersion (Rao, 1965) procedures were performed on landmark data which had been transformed by either the least-square or robust fit method to a common size, average position and orientation. Mutivariate tests of usefulness of covariates and tests of additional information and principal components analysis were each carried out according to Rao (1965) using matrix algebraic implementations of the techniques written by JGK in STSC APL. The Phylip package of tree building algorithms (Felsenstein, ) were used to compute UPGMA and search for the best Fitch-Margoliash (F-M) tree. An interactive APL F-M tree building program was written to facilitate exploring specific branching alternatives. The computer programs used in this work are available through the first author as MS-DOS executable programs or STSC APL workspaces.
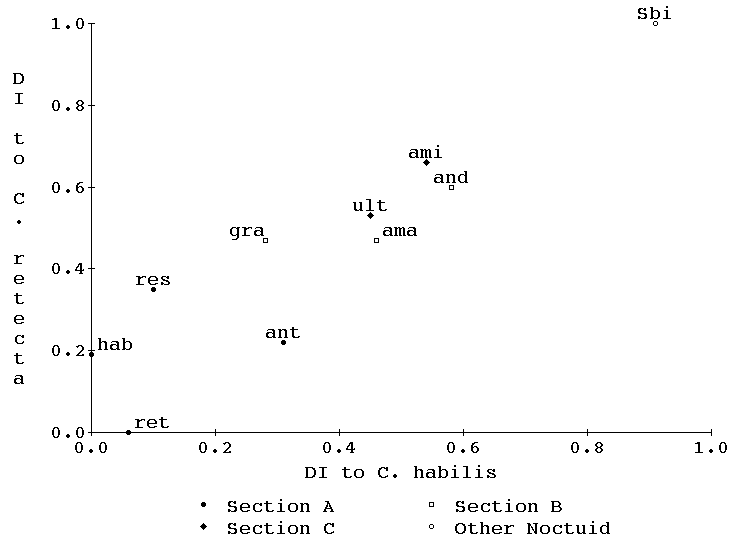
Yolk protein cross-reaction.-- The strongest cross-reactions of the two female-specific anti-yolk sera with non-homologous yolk extracts were obtained within the Catocala genus. The two outgroup species employed, Sunira bicolorago (Noctuidae; Cucullinae) and Amphipyra pyramidoides (Noctuidae; Acronyctinae), are outside the subfamily Catocalinae, and their yolk extracts cross-reacted modestly or not at all. The strength of cross-reactions within the Catocala (Fig. 4, 9 and 10) is consistent with previous divisions of the Catocala into Sections A, B and C based on foreleg spines (Hulst, 1884; Forbes, 1954) but does not agree entirely with the subgroupings (groups iv, v, vi, x, xi, xvii, xx), which are based primarily on visible forewing patterns (Forbes, 1954). In particular, Catocala dejecta (Group vi) was seen by anti-C. retecta-yolk (Fig. 10) to be further from C. retecta (also Group vi) than all other species in Section A (including Groups iv, v and vi). Within Section B, Catocala gracilis and Catocala andromedae, both Group xv, were placed relatively far apart according to their immunological cross-reactions to anti-yolks (Fig. 4), while most authorities have considered them very closely related (Barnes & McDunnough, 1917; Forbes, 1954). These early observations (Johnson, 1974) were published in abstract form (Kunkel et al., 1976) emphasizing the general agreement at the Section level without pointing out the cases where immunology and classical approaches strongly disagreed at the group level.
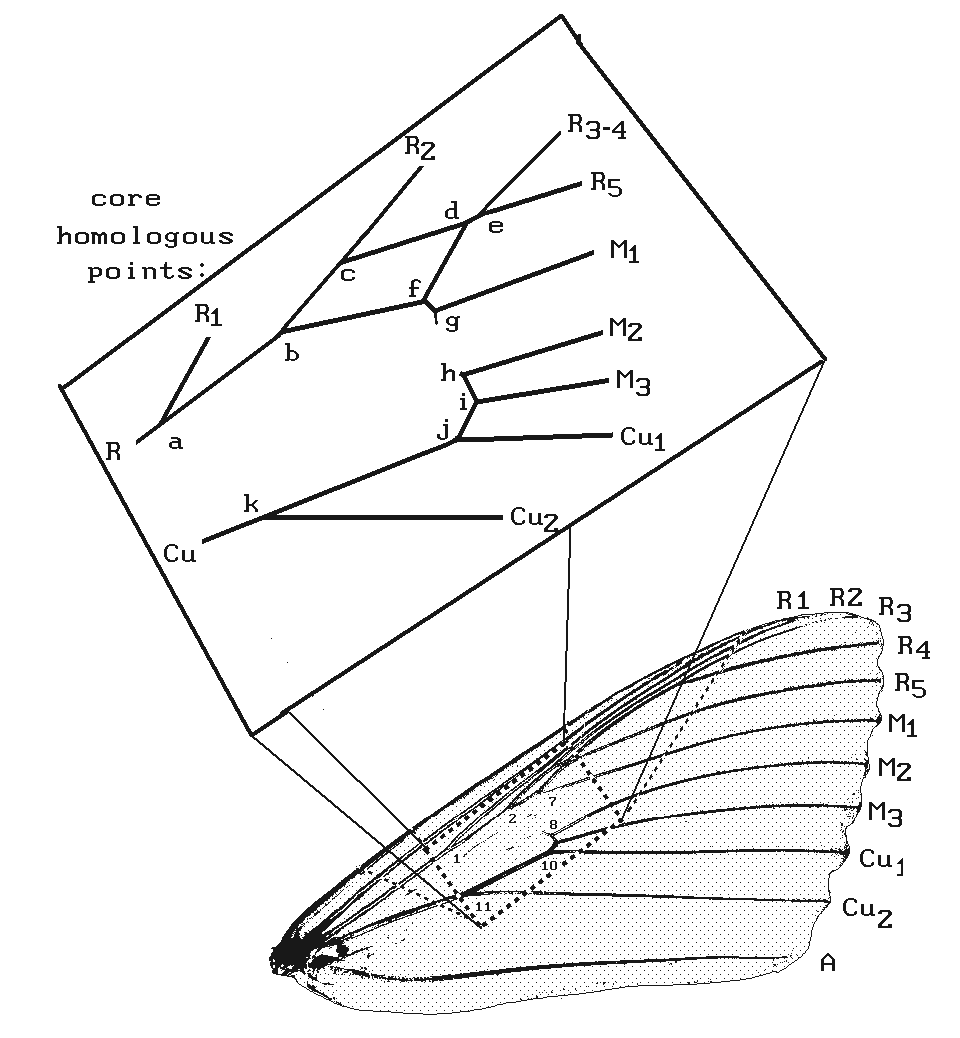
Choice of the wing landmarks.-- The choice of a central core of landmarks, avoiding the intersections of the veins with the wing margins (Fig. 5), was based on the general assumption that wing shape varies substantially with differing modes of flight behavior (Wootton, 1992) and that this variation has been selected for during evolution. We assert then that using the marginal landmarks would have confounded Darwinian selection for differences in shape with non-Darwinian neutral changes. A rigorous test of this assertion has not yet been carried out.
High intraspecific variability of wing morphology.-- While attempting to collect landmark data from a large array of Catocala wings, it became obvious that certain landmarks were variable in position even within species. In particular the insertions of radial branches R1, R2 and R3 into the main radial trunk, landmarks 1 2 and 3, were very variable. On an anecdotal level, when the right and left wings of an individual exhibited a strongly abnormal position, they did so as a pair, giving one the impression that the positions of the veins are symmetrical and thus not embryonic accidents (such accidents are often unilateral and not likely to be genetically controlled). Examples are given in Fig. 6 which illustrate three of seven C. epione specimens whose forewings exhibited obviously abnormal R2 behavior. In one, the R2 defined by origin originates in its usual position, based on other Catocala and our four 'normal' C. epione specimens, but crosses over R1 (and nominally becomes R1 at the margin of the wing.). In another of the extreme instances (Fig. 6A,B), R2 originates very late from the wing vein trunk (Fig. 6A but out of range in Fig. 6B). Roughly the same behavior is exhibited by the contralateral wing for each of the instances figured. In all instances the overall outline and shape of the wings are highly conserved. Thus the positions of the core wing landmarks appear to be highly conserved within individuals (comparing right with left wing) of a species but not between individuals of a species.
To demonstrate this phenomenon quantitatively, we utilized an F1 generation of 22 adults of a Catocala relicta mating which were raised on foliage in the laboratory (Sargent, 1972). An average model of the combined data from left and right wings was computed. Fig. 7 illustrates the 95% confidence ellipses for the 11 core landmarks of this wing population. It is clear that landmarks 1, 2 and 3, corresponding to the insertion sites of R1, R2 and R3, exhibit elongate confidence ellipses indicating their variable insertion points along the length of their originating trunk. Analysis of dispersion suggests that there is no significant within-individual variability between right and left wings. This result suggests that contralateral landmark traits are likely to be phenetically or genetically regulated in concert. It also suggests that a single wing from each individual is most efficient to compute the species average landmark distribution. The variability between individuals within a species, particularly in the position of landmarks 1, 2 and 3, argues that an average would be necessary to characterize a species venation pattern. Individuals of the same species, C. relicta, from widely different regions of the USA showed similar variability, but their average landmark position was quite close to the F1 generation distribution displayed.
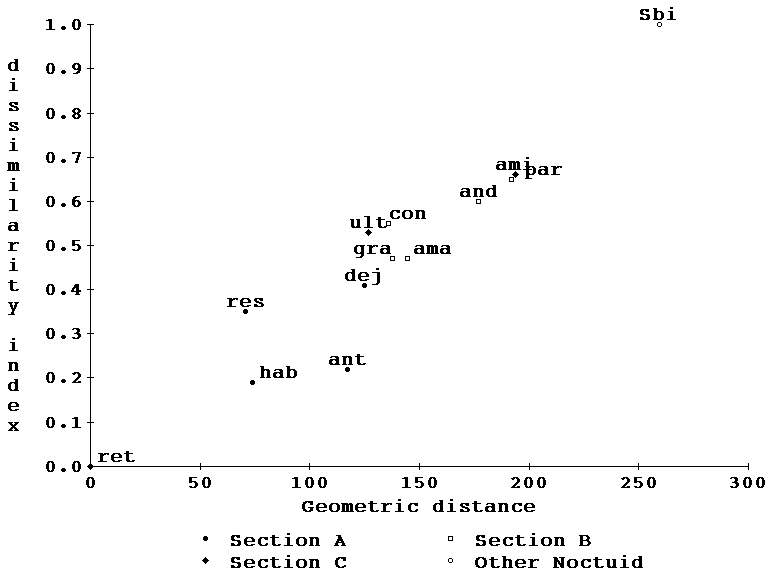
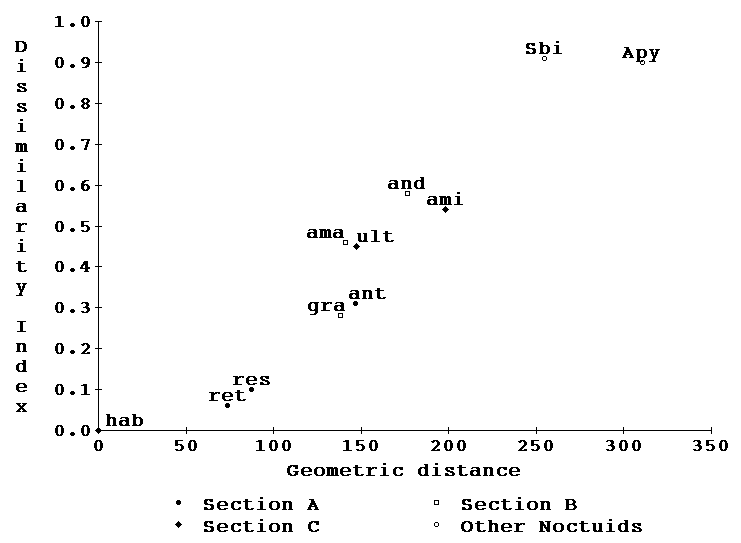
Interspecific comparison of wing morphology.-- Landmark averages were computed for all the species for which we had immunological data in the interest of subsequently comparing the morphological differences that correspond to the measured immunological distances. Catocala habilis and C. retecta were separately used as models and compared to all other species as in the immunological study. Fig. 8 is a plot of the differences measured in the same format as Fig. 4, and again C. dejecta plots distantly from C. retecta and C. gracilis plots remotely from C. andromedae.
Morphology vs immunology.-- Figs 4 and 8 data may now be combined into two additional figures, one comparing immunological and morphological distances of species from C. retecta (Fig. 9) and the other comparing similar axes relative to C. habilis (Fig. 10). Note that both the immunology and planar geometry agree on the remoteness of both C. retecta/C. dejecta and C. gracilis/C. andromedae. Of great interest is the fact that in both figures (9 and 10) the morphological and immunological distance measures form a joint straight line with both measures increasing by-and-large monotonically relative to the other. This high correlation suggests that both phenomena are relatively neutral to selectional pressures since neither shows any curvilinear departures from the other.
Phenetic tree building.-- Unlike the immunological study which was limited by a lack of antibodies to the yolks of all species of interest, the wing geometry approach allows each species' average to be used as the reference model. The use of C. retecta as a reference is depicted in Fig. 11, in which the residual vectoral differences in the positions of landmarks of each species fit to C. retecta are clustered emanating from a single origin. The average residual vector length, avg, and the root-mean-square residual vector length, rms, are both given as examples of how these metrics increase as the affinities of the species diverge. Thus a complete difference matrix, with each taxon serving as a reference, can be computed in a straightforward manner, Table II. Line four of Table II is derived from the rms metric illustrated in Fig. 11. In order to make the reciprocal tests close to equivalent, the size of each reference was isometrically adjusted to the size of C. retecta. The resultant matrix retains a degree of non-reciprocity since the method of least-square fitting, including scaling, rotation and translation, is not inherently reciprocal.
The PHYLIP algorithms, NEIGHBOR (UPGMA option) and FITCH, were applied to the distance matrices. The UPGMA option was used to establish a best branching sequence and then FITCH was applied to establish branch lengths as well as check the likelihoods of alternate branching patterns. Four criteria of goodness-of-fit of the resultant F-M trees were calculated. (1) The percent standard deviation, SD (Fitch and Margoliash, 1967), (2) the F value, a percentage difference in absolute branch lengths (Prager and Wilson, 1976) (3) n-, the number of negative branch lengths in the tree (Prager and Wilson, 1976), and (4) a newly devised chi-square statistic, using the observed pxp matrix of input distance matrix (Oij) and the expected pxp matrix of output distances (Eij) calculated from the sum of limb lengths of a tree fit by the Fitch-Margoliash algorithm. These were combined using the chi-square statistic,
Neutral genes are expected to diverge from one another by a process akin to diffusion. In such situations, as in diffusion, any change has a small likelihood of reversing itself by chance alone, so that the dominant overall result is divergence. A random walk brings one back to the origin in a small number of cases but more rarely as time progresses. Selection can either speed up or slow down this process of change if the trait is not, or ceases to be, neutral in a particular evolutionary lineage.
Both yolk proteins and the core landmarks of wing venation can be conceptualized as neutral traits. In both we are dealing with many details that might change by small increments without adversely affecting function. In the yolk protein precursor, vitellogenin, all the amino acids which are not directly involved in the few conserved properties of the protein are liable to change and such changes will on average tend toward the average protein composition based on the frequency of codon usage (King and Jukes, 1969). Changes in these amino acids decrease the number of immunological determinants and the avidity with which those determinants react to a heterologous polyclonal antiserum (Champion et al., 1974; Kunkel, 1988). A member of the vitellogenin family is the predominant yolk protein in the Lepidoptera. This family of proteins is conserved in general form throughout most of the Class Insecta (and indeed the animal kingdom). Vg is a large protein (subunit Mr ~200,000) which would be expected to have a large number of determinants, each of which would be subject to change and provide the basis for irreversibility. However, other abundant proteins which make up the balance yolk extracts in the Lepidoptera have been shown in a few instances to cross-react between closely related but not distantly related species (). The identity of all the antigens which contribute to the complete precipitin curves illustrated in Figure 3 has not been established. Several of the curves have indications of bimodality (also visible as double precipitin lines in the stained double diffusion gels) which are indications that the curves have a compound antigenic basis. This detraction from the simplicity of the immunological data and the substantial effort that would be required to resolve the details in each reference species is another reason that lends attraction to the alternate neutral-morphology approach.
In the core landmarks of wings, the two dimensions of freedom in which each landmark moves provide substantial freedom for that landmark and, in addition, the simultaneous random walks of the several landmarks provide a similar character of irreversibility to the divergence of landmark geometry. The statistical irreversibility of yolk protein and wing landmark evolution is an expression of Dollo's law which is a useful property of an evolutionary metric. While a pattern of multiple traits that has a selective advantage may be selected for, and that process may be reversed by selection against those traits, a pattern of multiple neutral traits once changed has a much smaller probability of being reversed. This beneficial property of a neutral metric of evolution is more precisely a result of the non-Darwinian forces of change, which have been intensively discussed for protein composition by King and Jukes (1969). There may be additional factors unique to neutral morphological traits which must be explored.
The rapidity of yolk proteins' evolutionary loss of immunological cross-reaction is an indication of its neutrality. The parallelism of yolk protein divergence and wing landmark divergence is both a confirmation of their mutual neutrality and an argument for their future value to an understanding of evolution and phylogeny. Any uneven Darwinian directedness of either trait's evolution would invariably have led to a curvilinear relation between their divergences, particularly since they are totally unrelated traits.
The use of yolk proteins as a neutral trait has been argued for before, but this is the first instance in which its neutrality has been supported by corroborating evidence. The size and the complexity of these protein molecules has interfered with their use for comparative phylogeny, but slow advances are being made. It should be recognized that the vitellogenin gene is homologous in birds, amphibians, insects and C. elegans (Spieth et al., 1985; Heine and Blumenthal, 1986; Wyatt, 1988). The structure of the molecule, a ca. Mr 500,000 dimeric protein, has been conserved despite the rapid change which the protein's evolutionary neutrality has allowed. Vertebrates have added the phosvitin domain which is missing in C. elegans and insects; thus the comparison and function across phyla are not exactly identical.
One might expect that wing venation might exhibit some of the same phenomena. Some of the wing landmarks we have used in Catocala, which are present in all Noctuidae, do not have homologues in other Lepidopteran families. One could chose a reasonable set of wing landmarks that did cover a larger proportion of the Lepidoptera. Despite the fact that insect wings are likely all homologous (Kukalová-Peck, 1978, 1983), it would be difficult to pick a set of homologous core landmarks for all the winged insects. While the basal origins and apical intersections of the traditional wing veins (marginal, radial, median, cubital, anal) with the wing margin might be homologized more broadly across the winged insects, the core venation details have undergone many changes, perhaps due to their neutrality. Further application of wing vein landmarks will be necessary to confirm the generality of their usefulness. If enough wing landmarks could be shown to have similar neutral rates of change within one order such as the Lepidoptera, it might be useful to compare the rates of non-homologous traits in different orders to achieve the objective of a comparable metric in different orders.
Our application of planar geometry of wings to produce a phenetic tree can be compared to a recent treatment of some of the same taxa (Gall, 1984). In particular our Section A species were dealt with by Gall in three phenetic trees based on 47 male genital characters and 18 egg and last larval instar characters, respectively. In Gall's analysis, the placements of species by the two independent sets of traits, genital traits and egg plus last larval characters, were not concordant with each other in predicting relationships of Section A species. However, one of Gall's three approaches to phenetic distances was substantially identical to our tree based on wing planar geometry (Fig. 13). Specifically, the genital based tree (Fig. 6.9 of Gall, 1984) agrees with the remote placement of C. dejecta/C. retecta. This is particularly pleasing since genital features have played such an important historic role in understanding alpha-taxonomy of insect species relationships. Since the other approach (Fig. 6.7 of Gall, 1985) using genital traits is not concordant with either Gall's second genital approach or our wing venation approach, we may have introduced an independent view directing a choice of Gall's two genital alpha-taxonomic approaches.
Our particular approach to analyzing differences in landmark locations rather than wing dimensions is a departure from the traditional wing morphological approach. While the planar geometry of mosquito wings has been used as an example in describing a specific planar geometric technique (Rohlf and Slice, 1990), we are not aware of another instance in which it has been usefully applied to exploring the evolutionary implications of the differences measured. Wing measurements have been effectively used in the past, but often there is an apparent multiplication of information through defining numerous distances all based on combinations of distances between a relatively few landmarks, i.e. strut analysis. Thus from five landmarks, A, B, C, D, E, one derives the 10 struts or combinations of distances between them, AB, AC, AD, AE, BC, ... , DE. The 11 landmark traits examined in our study would then have been represented by 55 struts between landmarks. The purpose of this latter approach is to capture, a priori, all the potentially important variables of wing structure. While the landmark location information undoubtedly is retained within the strut length data, it is confounded with information about size and correlated parallel struts. Multivariate analysis is required to extract the important information from the strut data. Our approach, planar geometry of landmarks, instead focuses on the two-dimensional placement of landmarks themselves, not the distances between them, and, without losing or ignoring any structural information, maintains the original complexity of the multivariate data (size differences are retained in a scaling parameter which may be of use in some studies).
The results of our early immunological observations of protein differences (Kunkel et al., 1976) were considered by us to be somewhat equivocal with earlier morphologically based affinities and differences (Hulst, 1884; Barnes and McDunnough, 1917; Forbes, 1954). We were not in a position at that time to claim that our molecular data were a better measure of phenetic difference than any other set of traits. An independent view of the relationships of these species was necessary to resolve the conflict. The introduction of planar geometric methods of studying landmark data (Siegel, 1981), and particularly our argument that a core set would behave neutrally, was seen as an attractive morphological approach particularly for wing venation (Rohlf and Slice, 1990). It allows a single metric on the distortion of landmark positions to represent the morphological distance between two taxa. It also allows multivariate analysis of the planar landmarks which have been rotated and isometrically scaled to a common model and orientation. Thus, our immunological approach has been confirmed by its essential agreement with an independent set of traits, the core wing landmarks, and additionally confirms a particular alpha-taxonomic approach using a traditional criterion of close affinity in insects, genital morphology.
While our work did not usefully overlap with the observations on variation in electrophoretic mobility for 13 proteins in five species of Catocala (Elliot and Johnson, 1979), our results concur with their observation that there can be substantial differences measured between supposedly closely related species.
It should be noted that there is a reluctance of some private collectors and museums to cooperate with our destructive approach (descaling) to analyzing the wing venation of their valuable specimens. Clearly this approach cannot be used to study the most important specimens of each species and variety, the holotypes and lectotypes in museum collections. We are acutely aware of this problem and are working on developing a non-destructive approach to recording venation landmarks.
It would be interesting to apply this wing geometry approach to Catocala in a more global perspective to see if this neutral trait approach would be of help in systematic placement of Catocala among its Ethiopian and Oriental relatives (Mitter and Silverfine, 1988).
We are hopeful that using these metrics on neutral traits will eventually allow us to place potentially directed evolutionary events (Darwinian traits), such as the appearance and population concordance of different chromatic and achromatic species of Catocala (Sargent and Owen, 1975; Sargent, 1978, 1981; MacLean, 1984), on an evolutionary (non-Darwinian) time scale that will help us to understand their origin and function. In a wider perspective, the potential also exists of linking molecular-based metrics such as mt-DNA, yolk protein, enzyme and serum storage protein evolution (Beverley and Wilson, 1984, 1985; Telfer and Kunkel, 1991) to the rates of morphological evolution in different families and orders of insects to see if morphological rates of change differ, as has been observed in vertebrates (Cherry et al., 1982; Wilson et al., 1984).
ACKNOWLEDGEMENTS
We pay homage to the late Allan C. Wilson who encouraged and stimulated our thinking about morphological vs molecular rates of change in diverse organisms. We are grateful to several private collectors, mentioned above, who donated their lepidopteran specimens to T. D. Sargent. We are indebted to several colleagues, particularly Ellen Prager and Larry Gall, who provided thoughtful criticisms of our manuscript. We thank the USDA-APHIS Methods Development Laboratory at Otis, MA, for aid and encouragement. This work was partially funded by an unrestricted grant from The EcoScience Corporation.
BARNES, W., AND J. MCDUNNOUGH. 1917. Check List of the Lepidoptera of Boreal America. Herald Press; Decatur, IL, 392pp.
BENJAMIN, D. C., J. A. BERZOFSKY, I.J. EAST, F. R. N. GURD, C. HANNUM, S. J. LEACH, E. MARGOLIASH, J. G. MICHAEL, A. MILLER, E. M. PRAGER, M. REICHLIN, E. E. SERCARZ, S. J. SMITH-GILL, P. E. TODD AND A. C. WILSON. 1984. The antigenic structure of proteins: A reappraisal. Ann Rev Immunol. 2:67-101.
BETTS, C. R., AND R. J. WOOTTON. 1988. Wing shape and flight behaviour in butterflies (Lepidoptera: Papilionoidea and Hesperioidea): a preliminary analysis. J Exp Biol. 138:271-288.
BEVERLEY, S. M., AND A. C. WILSON. 1984. Molecular evolution in Drosophila and the higher Diptera II. A time scale for fly evolution. J Mol Evol. 21: 1-13.
BEVERLEY, S. M., AND A. C. WILSON. 1985. Ancient origin for Hawaiian Drosophilinae inferred from protein comparisons. PNAS USA. 82:4753-4757.
BOYDEN, A. A. 1964. Perspectives in systematic serology. In: Taxonomic Biochemistry and Serology, Leone, C A, ed, Ronald Press Co; New York. pp. 75-99.
BRENNER, S., J. D. MURRAY, AND L. WOLPERT. 1981. Theories of biological pattern formation. Phil Trans R Soc. B295: 425-426.
BUTLER, J. E., AND C. A. LEONE. 1968. Determination of immunologic correspondence for taxonomic studies by densitometric scanning of antigen-antibody precipitates in agar-gel. Comp Bioch Physiol. 25: 417-426.
CARLSON, S. S., A. C. WILSON, AND R. D. MAXSON. 1978. Do albumin clocks run on time? Science. 200:1183-1185.
CHAMPION, A. B., E. M. PRAGER, D. WACHTER, AND A. C. WILSON. 1974. Microcomplement fixation. In: Biochemical and Immunological Taxonomy of Animals, Wright, C. A., ed, Academic Press; London and NY. pp. 397-416.
CHERRY, L. M., S. M. CASE, J. G. KUNKEL, J. S. WYLES, AND A. C. WILSON. 1982. Body shape metrics and organismal evolution. Evolution. 36: 914-933.
DICKERSON, R. E. 1972. The structure and history of an ancient protein. Scientific Amer. 226:58-72.
FELSENSTEIN, J. 1991. Allan Charles Wilson (1934-1991). Nature.{**** September:19 ****}.
FITCH, W. M., AND E. MARGOLIASH. 1967. Construction of phylogenetic trees. Science. 155:279-284.
FORBES, W. T. M. 1954. Lepidoptera of New York and Neighboring States. Noctuidae. Part III. Cornell University Experiment Station; Ithaca, New York. 309-341.
GALL, L. F. 1984. The evolutionary ecology of a species rich sympatric array of Catocala moths. PhD Dissertation. Yale University, New Haven, CN. Dis Abst 8509700, UMI, Ann Arbor, MI. 279pp.
HAGEDORN, H. H., AND J. G. KUNKEL. 1979. Vitellogenin and vitellin in insects. Ann Rev Entomol. 24: 475-505.
HEINE, U., AND T. BLUMENTHAL. 1986. Characterization of regions of the C. elegans X-chromosome containing vitellogenin genes. J Mol Biol. 188: 301-312.
HODGES, R. W. et al. 1983. Check list of the Lepidoptera of America North of Mexico. E. W.. Classey Ltd. and The Wedge Entomological Research Foundation, London, 284pp.
HULST, G. D. 1884. The genus Catocala. Bull Brooklyn Entomol Soc. 7:14-56.
JOHNSON, J. W., AND E. WALTER. 1978. Similarities and differences in forewing shape of six California Catocala species (Noctuidae). J Res Lepid. 17: 231-239.
JOHNSON, M. E. 1974. The evolution of yolk proteins in the underwing moths (Noctuidae, Catocala). Masters Degree Special Problem Report. University of Massachusetts, Amherst.
KING, T. H., AND J. L. JUKES. 1969. Non-darwinian evolution. Science. 164: 788-798.
KUKALOVA-PECK, J. 1978. Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record. J Morph. 156:53-126.
KUKALOVA-PECK, J. 1983. Origin of the insect wing and wing articulation from the arthropodan leg. Can J Zool. 61:1618-1669.
KUNKEL, J. G. 1975. Larval-specific serum protein in the order Dictyoptera- II. Antagonist effects of ecdysone and regeneration in the hemolymph of Blatta orientalis. Comp Biochem Physiol. 51B: 177-180.
KUNKEL, J. G. 1987. Analytical immunological techniques. in IMMUNOLOGICAL TECHNIQUES: ARTHROPODS ed. by LI Gilbert & TA Miller. Springer Verlag, NY. pp 1-41.
KUNKEL, J. G., M. JOHNSON, W. HAGGERTY, AND T. D. SARGENT. 1976. Conservation of an active site for oocyte recognition in rapidly evolving vitellogenins. Amer Zool. 16: abstract.
KUNKEL, J. G., AND J. H. NORDIN. 1985. Yolk proteins. Comprehensive Insect Physiology Biochemistry Pharmacology. 1: 83-111.
LEWIN, R. 1988. Molecular clocks turn a quarter century. Science. 239: 561-563.
MACLEAN, D. B. 1984. Evaluation of population attributes and life history traits of Catocala (Lepidoptera: Noctuidae) by means of multivariate analysis. Amer Mid Nat 112:67-75.
PRAGER, E. M. and A. C. WILSON. 1976. Congruency of phylogenies derived from different proteins. J. Mol. Evol. 9:45-57.
RADINSKY, L. 1978. Do albumin clocks run on time? Science. 200:1181-1183.
RAO, C. R. 1965. Linear Statistical Inference and Its Applications. John Wiley; New York, NY. 522.
ROHLF, F. J., AND D. SLICE. 1990. Extension of the Procrustes method for the optimal superimposition of landmarks. Syst Zool. 39:40-59.
SARGENT, T. D. 1969. A suggestion regarding hindwing diversity among moths of the genus Catocala (Noctuidae). J Lepid Soc. 23:261-264.
SARGENT, T. D. 1972. Studies on the Catocala (Noctuidae) of southern New England. III. Mating results with Catocala relicta Walker. J Lepid Soc. 26:94-104.
SARGENT, T. D. 1978. On the maintenance of stability in hindwing diversity among moths of the genus Catocala (Lepidoptera: Noctuidae). Evolution. 32:424-434.
SARGENT, T. D. 1981. On the achromatic Catocala. J Lepidop Soc. 35:81-93.
SARGENT, T. D., AND D. F. OWEN. 1975. Apparent stability in hindwing diversity in samples of moths of varying species composition. Oikos. 26:205-210.
SIEGEL, A. F. 1981. Geometric data analysis: an interactive graphics program for shape comparison. Modern Data Analysis, R L Launer and A F Siegel eds Academic Press. 103-122.
SIEGEL, A. F. 1982. Robust regression using repeated medians. Biometrika. 69: 242-244.
SIEGEL, A. F., AND R. H. BENSON. 1982. A robust comparison of biological shapes. Biometrics. 38:341-350.
SOKAL, R. R. 1961. Distance as a measure of taxonomic similarity. Systematic Zool. 10:70-79.
SOKAL, R. R., AND P. H. A. SNEATH. 1963. The Principles of Numerical Taxonomy. W H Freeman and Co; San Francisco, CA, 359pp.
SPIETH, J., K. DENISON, S. KIRTLAND, AND J. CANE. 1985. The C. elegans vitelogenin genes: short sequence repeats in the promoter regions and homology to vertebrate genes. Nucleic Acids Res. 13: 5283-5295.
TELFER, W. H., AND J. G. KUNKEL. 1991. Function and evolution of the hexameric storage proteins. Ann Rev Ent. 36: 205-228.
TELFER, W. H., AND C. M. WILLIAMS. 1953. Immunological studies of insect metamorphosis. Qualitative and quantitative description of the blood antigens of the Cecropia silkworm. J Gen Physiol. 36:389-413.
TILYARD, R. J. 1919. The Panorpoid complex. Part 3: the wing venation. Linn Soc NSW. 44:533-718.
WILSON, A. C., S. S. CARLSON, AND T. J. WHITE. 1977. Biochemical evolution. Ann Rev Biochem. 46: 573-639.
WILSON, A. C., J. G. KUNKEL, AND J. S. WYLES. 1984. Morphological distance: an encounter between two perspectives in evolutionary biology. Evolution. 38: 1156-1159.
WILSON, A. C., H. OCHMAN, AND E. M. PRAGER. 1987. Molecular time scale for evolution. Trends Genetics. 3: 241-247.
WOOD, B. 1983. Bones, teeth and molecular clocks. Review of New Interpretations of Ape and Human Ancestry. RL Ciochon & RS Corruccini, eds. Nature. 306: 140.
WOOTTON, R. J. 1990. The mechanical design of insect wings. Sci Amer. 263:114-120.
WOOTTON, R. J. 1992. Functional morphology of insect wings. Ann Rev Entomol. 37:113-140.
WYATT, G. R. 1988. Vitellogenin synthesis and the analysis of juvenile
hormone action in locust fat body. Can J Zool. 66: 2600-2610.
Table I. Enumeration (n) of Noctuid specimens used in study of wing venation. Identified by Genus, groups of Forbes (1954), species, (abr) abbreviation used in tables and figures, sex, collector, place and year. Sex was categorized as male (m), female (f), or unknown (?). Collectors included (FV) F. Vaughn, (GF) G. Forbes, (IL) I. Leone, (JGK) J. G. Kunkel, (TDS) T. D. Sargent. Places included (Amh-MA) Amherst, MA; (Brf-CT) Branford, CT; (Cry-IL) Carey, IL; (LaPl-CO) LaPlace, CO; (Lev-MA) Leverett, MA; (Pel-MA) Pelham, MA; (WHa-MA) W. Hatfield, MA.
Genus grp species abr n-sex Col Place Year Catocala iii epione epi 3m FV Amh-MA 1979 iii epione 1f JGK Amh-MA 1991 iii epione 1f,1m TDS Lev-MA iii epione 1f TDS Lev-MA 1971 Catocala iv antinympha ant 2?,2f,3m FV Amh-MA 1979 iv antinympha 1f FV Amh-MA 1970 iv antinympha 1m JGK Amh-MA 1991 Catocala v habilis hab 9m FV Amh-MA 1979 v residua res 1?,4m FV Amh-MA 1979 v residua 3m TDS Lev-MA 1970,8,8 Catocala vi dejecta dej 4m TDS Lev-MA 1970,1,2,3 vi palaeogama pal 7m FV Amh-MA 1979 vi retecta ret 2f,4m FV Amh-MA 1979 Catocala x parta par 3f TDS 1979 x relicta rel m FV Amh-MA 1979 x relicta 2f,1m GF LaPl-CO 1971 x relicta 1,10,1f TDS Lev-MA 1969,70,1 x relicta 1,11m TDS Lev-MA 1969,70 Catocala xi amatrix ama 3m CK WHa-MA 1969 xi amatrix 1m IL Cry-IL 1979 xi concumbens con 1m TDS Lev-MA 1971 xi concumbens 6m FV Amh-MA 1979 Catocala xv andromedae and 3?,2f,5m FV Amh-MA 1979 xv andromedae 1f TDS Amh-MA 1969 xv gracilis gra 1f,9m FV Amh-MA 1979 Catocala xvii ultronia ult 1f FV Amh-MA 1979 xvii ultronia 9m FV Amh-MA 1979 xvii ultronia ? JGK Amh-MA 1992 xvii ultronia ? JGK Amh-MA 1992 Catocala xx amica ami 1?,9m FV Amh-MA 1979 Amphipyra pyramidoides Apy 1m TDS Pel-MA 1964 pyramidoides 1m TDS Lev-MA 1976 Sunira bicolorago Sbi 2,1m TDS Brf-CT 1969,82 bicolorago 2? JGK Amh-MA 1992
Table II. Root-mean-square observed difference matrix of species (Sp) compared to a reference species (Ref) based on residual distances between landmarks after least-square fitting of species Sp to Ref. Species abbreviations as defined in Table I.
Ref\Sp: ant hab res ret dej par con ama and gra ult ami Sbi Apy ant 0 145 98 109 106 247 150 169 137 108 168 190 296 332 hab 145 0 89 76 115 174 125 142 180 130 146 200 282 317 res 98 89 0 71 90 215 111 124 144 107 119 173 257 340 ret 109 76 71 0 128 195 134 143 169 121 121 188 293 317 dej 106 115 90 128 0 247 146 140 151 135 190 218 243 358 par 246 173 214 194 247 0 173 240 238 184 173 166 386 326 con 150 125 111 133 146 174 0 170 117 86 102 133 269 281 ama 169 142 124 143 140 242 170 0 167 142 150 212 264 379 and 137 179 143 168 151 238 117 167 0 76 145 149 280 282 gra 108 130 107 121 135 185 86 142 76 0 105 115 298 289 ult 167 145 118 121 190 173 102 150 145 105 0 119 303 308 ami 189 199 172 187 218 166 133 211 149 115 119 0 362 323 Sbi 293 279 254 290 240 383 267 262 278 295 300 359 0 456 Apy 328 313 336 313 354 322 278 375 279 285 304 320 455 0
Figure Legends
Figure 1. Titration of precipitin reaction in gel with simultaneous dilution of antiserum and antigen. This curve allows prediction of expected precipitin in the case where an increasing antigen titer is desired but not possible. In such cases the antigen can be held constant and the antiserum is diluted. This curve estimates the factor which must be multiplied times the observed precipitin to obtain the expected precipitin at higher antigen doses.
Figure 2. Precipitin reaction in gel of Catocala habilis yolk with anti-C. habilis-yolk. Spectrophotometric scan at A550 across Thiazine Red R stained precipitin bands for Ag/Ab ratios of 1:8 to 16:1.
Figure 3. Sample full precipitin curves for (A) anti-C.retecta-yolk and (B) anti-C. habilis-yolk. Units of precipitate were obtained from the planimetric areas under curves such as figure 2. Averages of 6 curves for each Ag/Ab ratio are plotted vs Ag/Ab ratio as one full precipitin curve. The area under a full precipitin curves is computed by summing the non zero mean reactions. Variances of the grand mean are computed from the sum of the variances of the component means. The mean area under the homologous full precipitin curve, H, is used as reference and the precipitin dissimilarity coefficient, D, of a heterologous yolk precipitin curve, O, is measured as a proportional departure from this reference,
Figure 4. Precipitin dissimilarity to C. retecta is plotted against precipitin dissimilarity to C. habilis. The members of Section A, B and C are represented by filled circles, open boxes and filled diamonds respectively. The species outside the Catocalinae, Sunira bicolorago, is represented by an open circle.
Figure 5. Core landmarks in relation to the entire wing in C. retecta. A stick model of the venation is given in a blowup along with a verbal definition of each landmark.
Figure 6. Anecdotal anomalies in core landmark 2 for three C. epione forewings. Contralateral wings in the same specimens were highly correlated.
Figure 7. Scatter diagram of landmarks of individual specimen wings of an F1 generation of C. retecta plotted about a least-square average model for that species. A 95% confidence ellipse is drawn about each mean. Note that landmark 2 is particularly variable in the long dimension of the wing. This identifies the insertion of vein R2 to be particularly variable.
Figure 8. Geometric distances relative to C. retecta is plotted against geometric distances relative to C. habilis. Symbols as in figure 4.
Figures 9, 10. A synthesis of figure 4 and 8 data allows the geometric distances to be compared with precipitin dissimilarity. Symbols as in figure 4.
Figure 11. Planar geometric comparisons of species to C. retecta. Average landmark coordinates for 13 test species were fit to the reference species landmarks by the least-square approach of Siegel (1981). A. The positions of average core landmarks with connecting veins diagrammatically included is presented for the reference species, C. retecta, only. The residual vectors between C. retecta and A. pyramidoides are shown in situ and after 10 fold magnification collected about a single origin. B. The residual vectors, magnified by 10x, are given for all the test species in our immunological study, ordered into Catocala Sections A (the section of the reference species itself), B and C and other noctuids. The clustered vectors give a feeling for both the direction and magnitude of distortion of landmarks during evolution. The metrics avg and rms calculated for each test species are included for each cluster. The rms metrics of this figure correspond to the row 4 entries in the Table II matrix.
Figure 12. Phenetic similarity tree based on the matrix of root mean square differences in Catocala wing landmarks, Table II. Sections (A-C) and groups (iv-xx) are described by Barnes and McDunnough (1918) and Forbes (1954) respectively. This tree is the best fit of 19 trees constructed using the Fitch-Margoliash algorithm. Goodness of fit was adjudged by minimizing the four statistics SD, F, n- and chi-square described in the text. Negative branches are marked with an asterisk.
Figure 13. Phenetic similarity trees of six Catocala Section A species based on planar geometry. Branching for the wing geometry trees was suggested by three alpha-taxonomic studies by L. Gall (1984). Planar geometric distances were computed as in Table II on six species from section A for which we had wing venation data and were also treated by Gall (1984) in his figures 6.7, 6.9, and 6.11. The F-M tree building algorithm was applied to Gall's output data derived from his dendrograms to provide three phenetic trees with their branching sequence. The three branching sequences of Gall (left of A, B and C) were separately forced on an F-M treebuilding analysis (right of A, B and C) of the matrix of wing planar geometric distances. A fourth, best, F-M tree (D) for the wing venation data was found both by using the branching suggested by both the UPGMA method and the global search option in the FITCH algorithm. The best F-M tree is closest to Gall's male genitalic approach evinced in his Fig. 6.9 tree.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
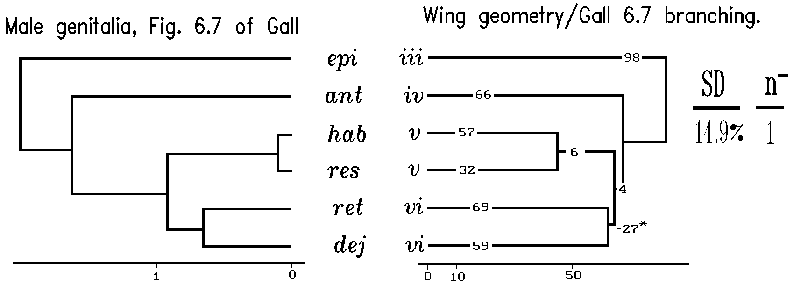{kind=link}
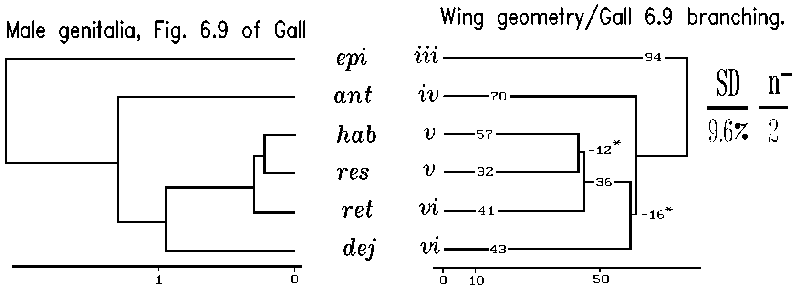{kind=link}
{kind=link}