Root growth: Balancing division and elongation
|||
N. B. Some of the text here has been edited down from Baskin 2013 WIREs Dev. Biol. 2:65-73, which is a useful resource if you want to read more about root growth kinematics. pdf: Baskin 2013
Animals are masters of motility; plants are masters of growth. Reacting to salient features of the environment whether biotic or abiotic, an animal might walk or slither, whereas a plant might grow thinner leaves or longer stems. Animals are typically stuck with a body from birth and acclimation is limited by physiology. In contrast, plants make organs continually throughout their lives, an iterative output allowing them to sidestep some physiological
constraints by virtue of development. In addition to its adaptive role for the plant, this incessant growth endows Earth with gigatons of biomass, sustains food chains, and colors our planet green.
Yay roots!
Because of their linear organization and radial symmetry roots make ideal subjects for investigating growth. In addition, unlike leaves, roots usually grow indeterminately, meaning that there is no programmed root length, and because the root grows continuously, its growth zone sampled at any time will contain the entire developmental sequence, from early to late.
As part of an acclimation response, it is common for the growth rate of a given root to increase or decrease. But which processes are changing? Root growth includes component processes, any or all of which, in principle, could change during acclimation. Surprisingly, some of these processes appear to be flexible and vary during acclimation, but others tend to be fixed. Recognizing this regularity helps us to zero in on the specific processes relevant for acclimation and also highlights the potential relevance of exceptions.
Kinematic framework
A powerful mathematical framework was applied to root growth as far back the 1950s. The approach is in general termed kinematic because it is in essence concerned with the movement of particles within the root and accounts for their motion with concepts from hydrodynamics. A formalism based on fluid flow applies well to the root growth zone (and to any plant organ) because the growth zone is continuous physically and expansion at any position causes surrounding tissue to move.
Because the mature part of the root is immobilized, expansion pushes the growth zone downward, through the soil. However, the zone does not move as a unit; instead the rate of motion (i.e., speed) at any point depends on the amount of expanding material between that point and the mature, non-growing part of the root. The speed goes from zero in the non growing region up to a maximum at the very tip of the root. The speed profile is thus a fundamental parameter containing information about the underlying expansion behavior within the root.
One aspect of research in the Baskin laboratory has been, and continues to be, to develop automated methods for quantifying the speed profile. These are described here. With a speed profile in hand, the span of the growth zone is delimited. In addition, the derivative of the speed profile gives the profile of elemental elongation rate, reflecting the local intensity of the expansion
What about cell division?
Happily, cell division can be characterized kinematically. In addition to the speed profile, this characterization needs the profile of cell length within the growth zone. We use cortex cells because they are easy to measure. At a given point in the root, the product of speed and cell density tells us how many cells are moving past that point per unit time. This parameter is called cell flux. If two nearby points have the same cell flux then no cells were produced between them; whereas, if the two points have different cell flux, then cells produced (or in principle destroyed) between them. Formalizing this relation mathematically allows rates of cell division to be calculated within the meristem, and also the location to be found where cell division stops (the base of the meristem).
Current projects
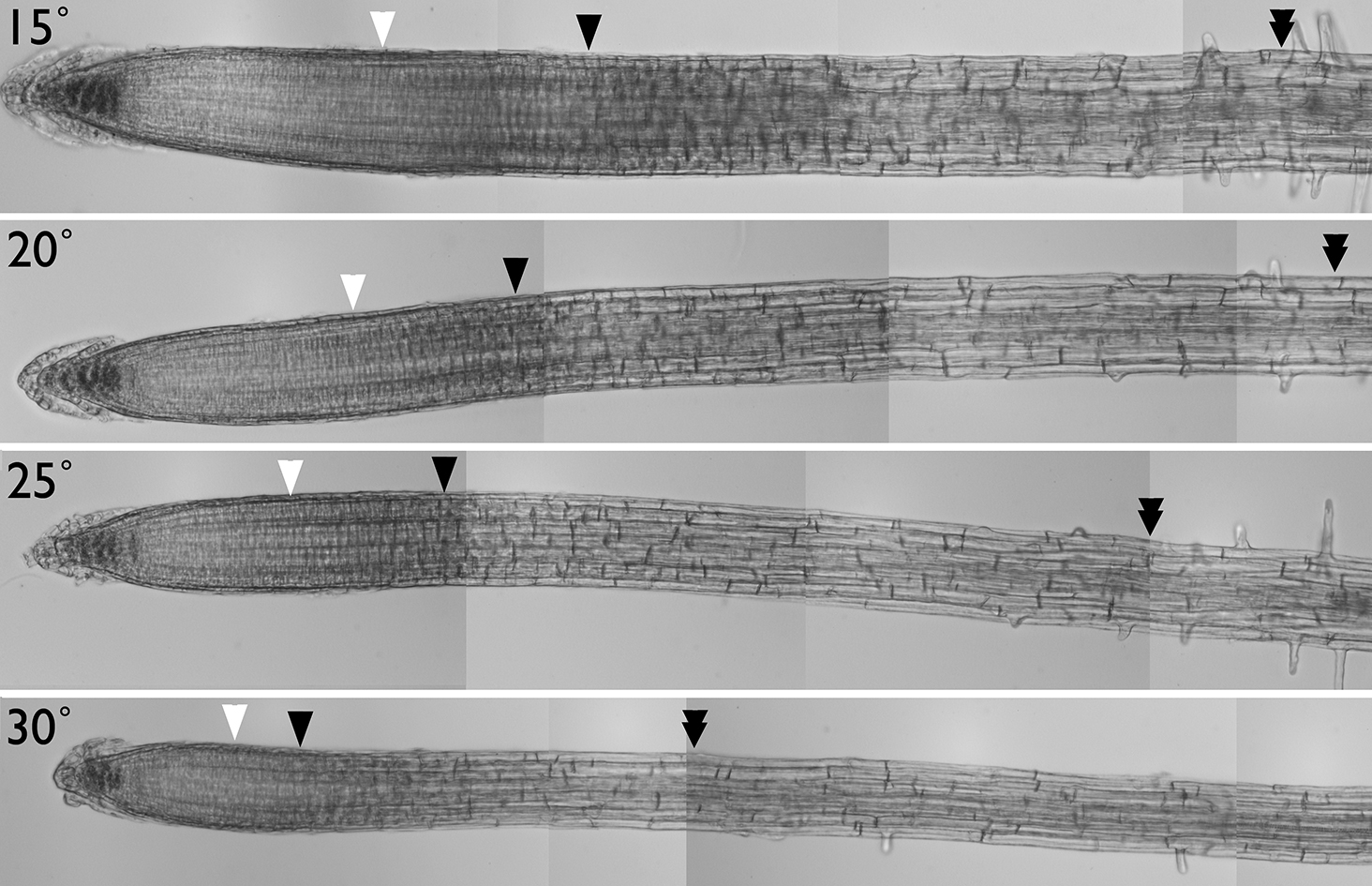
We are using the kinematic framework to investigate two issues. The first is the response of the root to ambient temperature. By ambient, I mean temperatures within a range where the root can more or less successfully acclimate, as distinct from a stress where the root has difficulty acclimating and goes into a special protection/survival regime. So far, we have found that, between 15 to 25 degrees C the total rate of cell production is temperature invariant. We are attempting to elucidate how this occurs and why. Our first paper in this area is here.
The second issue is understanding the nature of the transition between the meristem and rapid elongation zone. For this work, we are characterizing the growth at high resolution over several hours and we have found an intriguing oscillation in the position where elongation rate accelerates.
|
Roots grown at different temperatures Images are marked to show relevant boundaries, as found for that root (white arrowhead: shootward boundary of the meristem; black arrowhead: rootward boundary of the zone of elongation; double arrowhead: boundary of the zone of elongation). Bar = 100 microns. Image from Yang et al. 2017.
| |
|
| |
 Back to top Back to top |
|